| 視覚器の発生 |
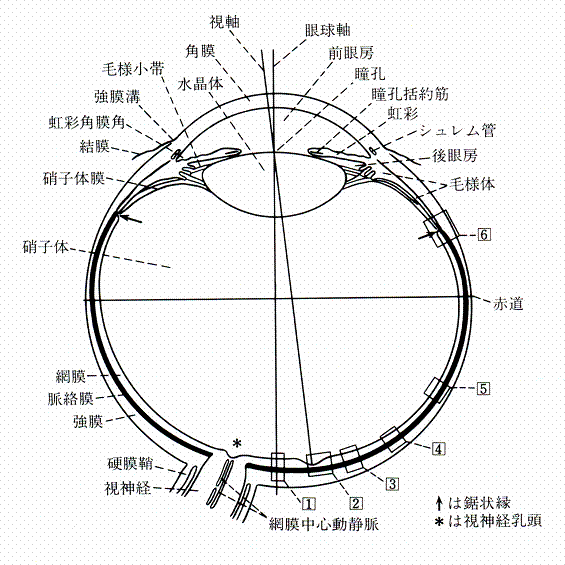 |
| 図1. 右の眼球の水平断面を上から見る |
 |
| 表1 眼球の構成 |
| 始めに |
| 視覚器は光の受容器である眼球と、その付属器である涙腺、涙道、眼瞼、外眼筋などから構成されている。 完成した眼球は、図1に示すような構造をしており、表1に示すように、いろいろの部分から成り立っている。これらの諸部の発生は複雑で、異なった種々の原基から発生する。すなわち、眼球の内膜は後に前脳になるべき脳の一部が膨出したものであり、水晶体は体表の外胚葉上皮が陥没することによって生じ、これらを外から包んでいる眼球血管膜と眼球繊維膜とは、間葉組織から形成される。また角膜上皮は体表の外胚葉上皮そのものである。 胎生第4週から第5週という発生の早期においては、胎児の体の各部を記載するには、方向用語を正確に決めておく必要がある。以下の記述においては、体の長軸方向を頭側(cranial)および尾側(caudal)方向とし、これに直角の方向を左右の外側(lateral)方向、腹背方向を腹側(ventral)および背側(dorsal)方向とする。また体の中軸部に近い方向を近位(proximal)、遠い方向を遠位(distal)とする。 |
| A. 眼杯および水晶体胞の形成 |
| 胎生第4週の初めごろ、神経板の頭側端部(将来前脳になるべき部分)において、左右の神経板の中央部に、頭尾方向に走る浅い溝が生じる。これが視覚器の発生の始まりで、この溝を視溝(optic sulcus)という。この溝は次第に深くなり、この溝を囲む神経板は外側方向に突出して、体表の外胚葉上皮の内面に接するようになる。 神経板が閉じて脳胞が成立する胎生第4週の終りごろになると、この突出は前脳胞の頭側端部の腹外側壁から外側方向に突出する球状の袋となる。これを眼胞(optic vesicle)といい、内部には前脳胞の内腔に続く腔が存在する。これを視室(optic vesicle)という。やがて前脳胞の背頭側端部が左右にむかって膨大して半球胞(終脳)を形成すると、もとの前脳胞は間脳胞と呼ばれるようになる。こうなると眼胞は間脳胞の頭側端部の腹外側壁から膨出することになる。 眼胞の遠位部は球状に拡大しながら外方に向かって突出していくが、近位部はあまり拡大せず、相対的に細い管状となり、眼胞茎(optic stalk)となる。 眼胞の遠位部が体表の外胚葉に接すると、この部の外胚葉上皮は増殖して、細胞の丈が高くなり、周囲の外胚葉の部分から明瞭に区別される円形の領域を形成する。この細胞の丈が高くなった領域から水晶体が形成されるので、この領域を水晶体板(lens placode)という。 やがて眼胞の遠位部は平たくなり、ついで近位方向に向かって陥没していく。その結果、始め1層の壁でできていた球状の眼胞は、内外二重の壁を持つ杯状の構造物となる。これを眼杯(optic cup)という。眼杯の内壁(これを眼杯内板という)の陥没が進むにつれて、はじめ広かった視室は次第に狭くなり、最終的には眼球内版が眼球外板に接着するので、視室は消失する。 眼杯内板が陥没を始めると、これに引かれるようにして水晶体板も陥没を始め、水晶体窩(lens pit)を形成し、さらに進むと、狭い開口部で体表に開く水晶体胞(lens vesicle)となり、胎生第5週の終りごろになると、その開口部が閉じ、さらに体表の外胚葉細胞との連絡も切れて、単層立方上皮で縁取られた、完全閉鎖性の球形の袋となり、眼杯内板が囲む腔の中に位置するようになる。 この時期においては、眼杯内板が囲む腔(これは将来の眼杯硝子体腔である)は、ほとんど完全に水晶体胞によって満たされており、眼杯内板と水晶体胞の間には、ごく少量の間葉細胞が介在するだけである。 眼杯および水晶体胞は、始めはこのように体の長軸に直角な面において、左右真横に向かって発生する。発生が進んで顔面が形成されると、眼杯と水晶体胞は次第に顔面の正中線に近づいてきて、前方を向くようになる。こうなってから後は、眼杯と水晶体胞については、光が入ってくる遠位側を前方、反対の近位側を後方と記述する。 水晶体胞は、上に述べたように、体表の外胚葉上皮に由来する1層の立方上皮で縁取られている。この上皮のうちで、体表に近い面(前面)を作っているものは、細胞の丈が高くならず、終生、単層立方上皮の状態を維持する。これを水晶体上皮(lens epithelium)という。これに対して、後面を作っている上皮細胞は、著しく細胞の丈が高くなり、前後方向に細長い繊維状の細胞となって、水晶体胞の内腔に隆起していき、胎生第7週の終りごろになると、その前端が前面の水晶体上皮の後面に接着して、水晶体胞の内腔を完全に閉鎖する。この後面を構成する繊維状の細胞を水晶体繊維(lens fibers)という。水晶体繊維が前面の水晶体上皮に移行する部位を、水晶体の赤道(lens equator)という。このようにして、水晶体の原型が成立する。 |
| B. 眼裂およびその閉鎖 |
| 眼杯内板の陥没は極めて特異な経過をとる。すなわち、この陥没は眼胞の中心部において同心円性に起こるのではなくて、眼胞の腹側縁に近い所を中心にして偏心性に起こる。したがって、陥没が進行すると眼杯の腹側縁には1つの切れ込み、ないし裂け目が生じる。この裂け目を眼杯裂という。この裂け目は眼杯の腹側縁から、眼杯茎の遠位部の腹側面にまで達しており、ここでは眼杯茎裂と呼ばれる。眼杯裂と眼杯茎裂とをあわせて眼裂または胎生裂(choroidal or foetal fissure)という。 その後の発育において、眼杯内板は網膜に、眼杯外板は網膜色素上皮層になるのであるが、眼杯内板のうちで、眼杯茎裂に直接接している腹側部以外の残りの大部分は、眼杯茎裂のところで眼杯茎の腹側壁に直接続くことになる。これによって、眼杯内板すなわち網膜に発生した神経細胞の軸索(視神経繊維)が、眼杯茎を経て間脳に達する道が確保される。このように、眼杯内板の陥没が眼胞の腹側縁に近いところを中心にして、偏心性に起こることは、眼杯内板と眼杯茎の直接の連絡を確保しつつ、二重壁の眼杯を形成していくための、実に巧妙な仕組みである。 胎生第5週の後半において胎生裂が成立すると、内頚動脈から枝別れして眼杯の基部に達している眼動脈から1本の枝が出て、胎生裂の中に進入する。この動脈は少量の間葉組織を伴って、胎生裂の中を前進し、さらに眼杯腔(将来の硝子体腔)の中軸部を通って水晶体胞の後面に達し、ここで水晶体胞を包む血管網をつくる。この動脈を硝子体動脈(hyaloid artery)という。 胎生裂は一時的なもので、硝子体動脈が胎生裂に入ると、間もなく、胎生裂を左右両側から囲んでいる眼杯および眼杯茎の自由縁が互いに癒着して胎生裂を閉ざす。こうなると、眼杯の内板と外板とは、それぞれ一続きとなり、眼杯は初めて完全な杯状となる。眼杯内板が眼杯外板に反転する部分は、前方に向かって正円形の孔を囲む。この孔が将来の瞳孔(pupil)である。 もし、何らかの原因で胎生裂の閉鎖が妨げられると、瞳孔から下方に伸びる裂け目が生じる。これをコロボーマ(coloboma)といい、その程度に従って、虹彩コロボーマ、脈絡膜コロボーマと、虹彩および脈絡膜コロボーマを区別するが、最も高度な場合には網膜も合一せず、瞳孔から視神経乳頭に達する裂け目が存在する例も観察されている。 硝子体動脈が硝子体腔に進入する部位から、眼杯内板の表面に沿って走り、眼杯内板に分布する枝が出る。胎生の後期になると、硝子体腔の中軸部を貫通して水晶体の後面に達していた、本来の硝子体動脈は退化して、消失し、眼杯内板、すなわち、網膜に分布するこの枝だけが残る。これを網膜中心動脈(central retinal artery)という。網膜を養う血管が網膜の内面から網膜の内部に進入することは、脳の血管が脳の外表面から実質内に進入しているのと同じである。硝子体に接する網膜の内面は脳の外表面である。 |
| C. 眼杯内板と眼杯外板の分化 |
| 眼杯の内板と外板は、はじめは共に多列円柱上皮で構築されている。外板は眼杯の拡大につれて次第に薄くなり、単層円柱ないし単層立方上皮となる。胎生第5週の後半になると、この細胞の胞体内に黒褐色のメラニン顆粒が多量に出現し、全体として網膜色素上皮層(retinal pigment epithelium)となる。これによって眼球原基は、ほとんど無色のこの時期の胎児において特に目立つ。 眼杯内板は盛んな細胞分裂によって厚くなり、さらに細胞に分化が起こって、光を感知する感覚細胞と、その興奮を脳に伝達する神経細胞とが生じ、全体として網膜(retina)となる。しかし、瞳孔縁に近い一定の範囲では、このような肥厚も細胞の分化も起こらず、最終的には単層立方上皮となり、その外側に接している網膜色素上皮層とともに、毛様体および虹彩の上皮性部を形成する。この部分を網膜盲部(pars caeca retinae)という。これに対して光を感知する残りの部分を網膜視部(pars optica retinae)という。 |
| D. 水晶体の形成 |
| 水晶体の原型が、上に述べた経過で成立すると、これに引き続いて水晶体の原型の赤道付近で水晶体上皮の増殖が始まり、新しい水晶体繊維が原型の外側に新しい層をなして、次々に付加されていく。新しい水晶体繊維は、前面では水晶体上皮の後面に接して前極に向かって伸長し、後面では水晶体の後面を後極に向かって伸びる。 1本の水晶体繊維の長さは、前極から後極までの長さの約半分であり、前面で前極に近いところから始まる繊維は、後面では後極から遠いところで終わる。前面における起始部が前極から遠ざかるにつれて、後面における終止部は後極に近づく。1つの層の内部では水晶体繊維の長さはほぼ同一であるから、水晶体の赤道上の向かい合う2点から出る水晶体繊維の会合点を連ねる腺は、水晶体の前面および後面を3等分する倒立Y字形(前面)、および正立Y字形(後面)の図形を描く。これを前および後水晶体星(anterior and posterior lens stars)という。 水晶体上皮が水晶体繊維に移行する赤道部より後方では、水晶体の後面は均質無構造の水晶体被膜(lens capsule)によって包まれている。この被膜は胎生第6月ごろから認められるようになる。 水晶体繊維の新生と付加は、生後も引き続いて行われ、20歳ごろになると一応終了する。しかし、水晶体繊維の付加は、きわめて緩やかながら、終生持続するものと信じられている。 |
| E. 水晶体血管被膜 |
| 水晶体は上述のように、体表の外胚葉上皮に由来する水晶体胞から形成され、間葉組織は全くその形成に関与しない。しかし、胎生期間中は、水晶体は血管網を含む疎な間葉組織によって包まれており、これから必要な栄養分を供給されている。この間葉性機膜を水晶体血管膜(vascular membrane of the lens)という。 水晶体血管膜の血管の大部分は硝子体動脈に由来する。この動脈は水晶体の後面に達すると、多数の小枝に分かれて、水晶体の後面を包む血管網を形成する。さらに一部は水晶体の赤道を越えて水晶体の前面に現れ、ここで水晶体の前面を包んでいる間葉組織の中の血管網と吻合する。 |
| F. 硝子体 |
| 眼杯が成立したばかりの時点では、水晶体胞は相対的に非常に大きくて、眼杯内板が囲む空間をほとんど完全に満たしており、眼杯内板と水晶体胞の間には、ごく少数の間葉細胞が介在しているに過ぎない。眼杯の成立経過から明らかなように、眼杯腔を囲む眼杯内板の表面(内面)は脳の外表面そのものであり、これが基底膜を介して間葉組織に接していることは、言うまでもない。 胎生第6週に入ると、眼杯が水晶体胞よりもずっと速やかに拡大していくので、眼杯内板と水晶体胞の間の隙間は急速に拡大して、広い腔となる。この腔を硝子体腔(hyaloid cavity)という。この硝子体腔の中軸部を硝子体動脈が、少量の間葉組織を伴って、後ろから前に向かって走る。硝子体腔はこうして拡大していくが、このさい間葉組織はあまり増加せず、拡大した硝子体腔はほとんど細胞を含まない無色透明なゼリー状物質で満たされる。この状態を第一次硝子体(primary vitreus)という。 胎生第9週以後になると、眼杯内板、すなわち、網膜の内面を裏打ちしている基底膜に終わる微細な繊維が出現して、第一次硝子体の周囲に無細胞性の網工を形成する。ついでこの網工は無色透明のゼリ状物質で埋められる。これを第二次硝子体という。発生が進行するにつれて、第二次硝子体は急速に増加し、第一次硝子体は次第に硝子体動脈の周囲の狭い管状の領域(硝子体管 hyaloids canal)に限局されるようになる。胎生の後期において、硝子体動脈が退化し、消失すると、その周囲の第一次硝子体も消失し、出生後には硝子体管は痕跡すら認められない。 硝子体の前縁では、硝子体の組織がやや密になっており、薄い硝子体膜として認められる。 |
| G. 眼杯の間葉性被膜 |
| 眼杯は成立の始めから、その全周を間葉組織によって包まれている。この間葉組織はやがて眼杯に直接接する疎な内層と、その外側を包む緻密な外層とに分かれる。内層は一般に血管に富むので、眼球血管膜と呼ばれ、網膜視部では脈絡膜(choroid)を、網膜毛様体部では毛様体の実質を、網膜虹彩部では虹彩支質(iris stroma)を形成する。外層は眼球繊維膜と呼ばれ、厚い緻密な繊維性結合組織となって、眼球の形を維持する強膜(sclera)と角膜固有質(corneal stroma)を形成する。 外層はさらにその外側を包む頭部の間葉組織に移行する。この頭部の間葉組織から眼球鞘や眼窩脂肪体が発生する。眼球を動かす外眼筋も、この間葉組織の内部に出現する。 |
| H. 脈絡膜 |
| 眼杯が成立すると、間も無く眼杯外板に密着して毛細血管網が形成される。これが脈絡膜の始まりである。胎生第8週になると、この脈絡膜原基の外側に間葉細胞が密集してきて、強膜の形成が始まる。この時期においては、脈絡膜はなお1層の毛細血管網だけでできている。胎生第4月になると、眼杯外板、すなわち、網膜色素上皮層に密着するこの毛細血管網(これを脈絡毛細血管板という)の外側に、やや太い血管の網が出現して、脈絡血管板を形成する。胎生の後半になると、これらの血管の間の隙間は、神経堤に由来する色素細胞によって埋められる。脈絡膜の血管は眼動脈から出る多数の小動脈に由来する。 網膜色素上皮層と脈絡毛細血管板の間には、弾性繊維を含む著明な基底膜が存在する。これをブルッフ膜(Bruch’s membrane)という。電子顕微鏡によると、この膜は、色素上皮細胞の基底板と、脈絡毛細血管板の毛細血管の色素上皮層に向かう側の基底板、および両者の間に介在する弾性繊維によって構成されている。この弾性繊維が出現するのは胎生第4月以後である。 |
| I. 角膜と強膜 |
| 胎生第5週において、水晶体が成立した時点においては、水晶体と体表の外胚葉上皮の間には、扁平な細胞がごく少数介在しているに過ぎない。胎生第6週になると、眼杯の周囲を包んで次第に増加してきた間葉細胞が、眼杯の前縁を越えて、外胚葉上皮と水晶体胞の間に進入してきて、眼杯の前縁および水晶体胞の前面と外胚葉上皮の間を、次第に密に埋める。 胎生第7週に入ると、この間葉組織のうちで、水晶体胞の前面に接する間葉細胞が上皮細胞様に変化して、著明な単層扁平上皮として、間葉細胞の後面を縁取るようになる。これを角膜内皮(corneal epithelium)という。 このような変化につれて、水晶体胞の前方を被っていた体表の外胚葉上皮も変化を始め、始めの単層立方上皮の状態から次第に多層化して、胎生第8週には3〜4層の細胞からなる重層扁平上皮となる。これが角膜上皮(corneal epithelium)である。角膜上皮と角膜内皮の間の間葉細胞は、次第に表面に平行な層を作って整然と配列し、細胞間には多量の膠原繊維が出現し、これらは極めて密に整然と配列して、層板状緻密結合組織となり、全体として厚い角膜固有質となる。 角膜固有質は眼杯の前縁のところで強膜に移行するが、強膜に膠原繊維が出現するのは、角膜におけるよりもずっと遅い。また強膜における膠原繊維の配列は、角膜固有質におけるほど規則正しくはなく、交織性緻密結合組織と層板状緻密結合組織とが混ざり合った状態である。 角膜内皮も、眼杯の前縁付近で虹彩の原基である間葉組織に移行する。この移行は突然で、単層扁平上皮が突然星型の間葉細胞に移行する。この部の間葉組織から、虹彩角膜角(前房隅角)における櫛状靭帯(pectinate ligament)が発生する。 |
| J. 虹彩と毛様体 |
| 胎生第8週を過ぎると、眼杯の拡大は著しく加速されるが、水晶体の増大は比較的緩やかであるので、水晶体は形成されたばかりの角膜から離れて、次第に後退して、再び眼杯の中に位置するようになる。このさい、角膜と水晶体の間に散在していた間葉細胞は大部分消失して、ここに細胞が存在しない空間が成立する。これが前眼房(anterior chamber)である。ただし、水晶体の前面に密着していた間葉細胞は消失せず、眼杯の前縁の前面を包む間葉組織の濃縮部に連なって、水晶体の前面を被う薄い膜を形成する。これを瞳孔膜(pupillary membrane)という。瞳孔膜には、やがて血管網が発生して、水晶体を前面から栄養する装置となる。 胎生第9週の終りごろから、眼杯外板が眼杯内板に反転する眼杯の前端の部分が、水晶体の赤道部を越えて、瞳孔膜を狭めながら、水晶体を前面から被うように発育していく。この新たに発育を始めた部分では、眼杯内板に上皮細胞の多層化が起こらず、内板はやがて単層立方上皮となり、その外側に接する眼杯外板と共に、虹彩(iris)および毛様体(ciliary body)の上皮性部を形成する。これが網膜盲部である。単層立方上皮である網膜盲部と、多層化した網膜視部の境界線が鋸状縁(ora serrata)である。 眼杯の前縁の外周を囲んで濃縮していた間葉組織は、この上皮性部の外側(そとがわ)に付随して伸びていき、虹彩支質および毛様体の実質の原基となる。虹彩および毛様体の形成と成長は、眼球の他の構造に比べると非常に緩やかで、胎生第6月においてもこれらの形成はなお微弱である。 網膜盲部のうちで、水晶体の前面を被うように発育していく部分が網膜虹彩部(pars iridica retinae)で、これが網膜色素上皮層に反転するところが瞳孔縁である。一方、網膜虹彩部と網膜視部の間の部分は、水晶体の赤道部を囲む部分であるが、この範囲では、網膜盲部は網膜色素上皮層とともに、眼球の内腔に突隆する多数の「ひだ」を形成する。この「ひだ」は瞳孔から四方に放散する線、すなわち、経線(meridian line)に沿って整然と配列しており、毛様体突起(ciliary process)と呼ばれる。この部分の網膜盲部を網膜毛様体部(pars ciliaris retinae)という。 眼杯外板である網膜色素上皮層と、眼杯内板である網膜虹彩部および網膜毛様体部とは、互いに視室に向かい合う自由表面どうしで密着している。したがって、網膜色素上皮層の外面と、眼杯の内腔に向う網膜虹彩部および網膜毛様体部の内面は、どちらも脳の外表面に相当し、著明な基底板によって裏打ちされている。 虹彩の部分では、上皮性部の前面を被う間葉組織によって虹彩支質が形成され、その内部に、瞳孔の中心から四方に放散するように、放射状に配列する多数の血管が発生する。胎生第6月の終りごろになると、脈絡膜の中を後方から前方に向って貫いてきた長後毛様体動脈が虹彩の基部に達し、ここに大虹彩動脈輪を形成して、虹彩への血液供給の基地となる。 虹彩支質は間葉細胞の増加によって次第に厚くなるが、繊維成分の発生はごく少量である。間葉細胞はやがてメラニン顆粒を獲得して色素細胞となるので、虹彩支質は次第に黒くなる。また、胎生第7月の終りごろになると、網膜虹彩部の細胞も、色素上皮層の細胞も、メラニン顆粒によって細胞体を埋め尽くされる。 虹彩の色素上皮層の細胞から2種類の平滑筋が発生する。第1は胎生第4月の経過中に、瞳孔縁に近い比較的狭い範囲の色素上皮細胞から生じるもので、これらは間も無く色素上皮層から離れて虹彩支質の中に埋まり、瞳孔の外周を取り巻いて輪状に配列し、全体として瞳孔括約筋(iris sphincter muscle)となる。第2は、瞳孔括約筋が発生した領域を除く、虹彩の全領域から生じるもので、全体として瞳孔散大筋(iris dilator muscle)を形成する。この領域の色素上皮細胞は、核よりも基底側、すなわち、虹彩支質に接する側の細胞質は、細長く伸びて平滑筋の性質を獲得し、放射状に配列して瞳孔散大筋となるのであるが、核よりも自由表面側、すなわち、網膜虹彩部に接する側の細胞質は色素上皮細胞としての性質を保っている。 このように、瞳孔括約筋と瞳孔散大筋は、神経板由来の外胚葉性上皮細胞から発生した特異な平滑筋である。 虹彩が水晶体の前面を被うように発育していくにつれて、はじめ水晶体の前面の全体を被っていた瞳孔膜は、次第に相対的に狭くなり、虹彩の瞳孔縁が囲む瞳孔を閉ざすのみとなる。この瞳孔膜は胎生第9月の経過中に退化して消失し、出生時にはその痕跡も認められない。もし、何らかの原因で、瞳孔膜の退化・消失が妨げられて、出生時に瞳孔膜の全部または一部が残存している場合には、これを瞳孔膜残遺という。 毛様体は、完成した眼球では、毛様体突起をそなえて眼球の内腔に隆起している毛様体ひだ部(pars plicata)と、その後方に続く平滑な毛様体扁平部(pars plana)の2部からできている。しかし、毛様体扁平部の発育は遅く、胎生第6月の終りにおいてもなお認められず、網膜視部と網膜盲部の境である鋸状縁は、毛様体筋のほぼ中央部の位置にある。胎生の後半において、眼球全体が拡大し、水晶体が次第に眼球の前方部に位置するようになると、水晶体の赤道部を囲む毛様体ひだ部も前方に進み、これに伴って、その後方に毛様体扁平部が認められるようになる。 毛様体の上皮性部の外側(そとがわ)を包んでいる間葉組織は、盛んに増殖して多数の平滑筋を生じ、全体として発育のよい毛様体筋(ciliary muscles)を形成する。最初に出現するのは経線繊維(meridional fibers)で、胎生第5月において既に識別される。これに対して、輪状繊維(circular fibers)と放射繊維(radial fibers)が認められるようになるのは胎生第7月以後であり、しかもこれらの筋が完成するのは出生後である。 毛様体の上皮性部の細胞のうちで、外側(そとがわ)の色素上皮層の細胞は非常に早く、胎生第5週の後半からメラニン顆粒を獲得するが、内側(うちがわ)の網膜毛様体部の細胞は、終生、無色透明の単層立方上皮の状態を維持する。この上皮細胞は水晶体を養い維持する眼房水(aqueous humor)を分泌する。この細胞の眼杯内腔に向かう面(これは基底面に他ならない)に接して、厚い著明な基底膜が存在し、ここに水晶体を支持する小帯繊維(zonular fibers)が付着して、Zinn小帯(毛様小帯)となる。この小帯繊維の発生機序については、今日なお明らかでない。 後眼房(posterior chamber)は、主として虹彩の後方にあった水晶体血管膜およびこれに関連していた間葉細胞が消失することによって成立する。胎生第9月において瞳孔膜が消失すると、後眼房は瞳孔を通して前眼房に連なる。前眼房および後眼房は、網膜毛様体部の細胞が分泌する眼房水によって満たされている。眼房水は後眼房から瞳孔を通って前眼房に入り、虹彩の前面および角膜の後面を潤したのち、前眼房隅角部の繊維柱帯のところからシュレム管を経て排出される。 |
| K. 網膜の組織発生 |
| 網膜視部は、これまで述べてきたことから明らかなように、脳の一部であり、その組織発生の過程は中枢神経系の他の部分におけるものと原則的には同一である。ただし、眼杯内板が陥没して眼杯硝子体腔を囲むようになり、はじめ脳の外表面であった面が眼杯の内面となるので、脳(中枢神経系)の組織発生における「うちがわ」から「そとがわ」に向う組織分化の方向が、網膜では逆に「そとがわ」(色素上皮層に向うがわ)から「うちがわ」(硝子体腔に向うがわ)に向うことになる。 網膜視部は、はじめは、数列の核が一様に並んだ多列円柱上皮でできている。やがて盛んな細胞分裂によって全体として厚くなり、眼裂が閉じる胎生第5週の後半になると、眼杯硝子体腔に面する「うちがわ」の部分に、核を含まない層が識別されるようになる。これは脳の発生における縁帯(marginal layer)に相当し、核の密集した「そとがわ」の層は胚芽層(matrix layer)に相当する。 胎生第6週の中頃になると、胚芽層から内方に向って細胞が遊出を始め、第7週の終りごろになると、胚芽層の「うちがわ」に明瞭な細胞層が形成される。この層は脳の発生における外套層(mantle layer)に相当し、幼若な神経細胞によって構築されており、内神経芽細胞層(inner neuroblastic layer)と呼ばれる。この層と胚芽層(網膜の発生においては外神経芽細胞層 outer neuroblastic layerと呼ばれる)の間には、狭い無核性の層が介在する。 内神経芽細胞層は、外神経芽細胞層から遊出する多数の幼若神経細胞の参加によって、次第に厚くなり、胎生第3月の終りごろになると内外の2層に分かれる。「うちがわ」の層は、これを構成する神経細胞の軸索(神経線維)が視神経繊維になるので、視神経細胞層(神経節細胞層 ganglion cell layer)と呼ばれる。この層の神経細胞の軸索は内方に進んで縁帯に入り、ここで方向を転じて眼杯の内面に平行に走って眼杯茎に向って進み、眼杯内板と眼杯茎の結合部を経て眼杯茎に入り、眼杯茎の中を後方に進んで脳(間脳)に達する。このようにして、神経細胞層の「うちがわ」の縁帯は、視神経繊維で埋められて視神経繊維層(optic nerve fiber layer)となる。また眼杯茎は視神経線維で満たされて視神経(optic nerve)となる。 「そとがわ」の層は、元来の内神経芽細胞層そのものであり、ここに残った幼若神経細胞は、この位置で成熟して、その大部分は小型の双極神経細胞となる。この層は、完成した網膜における内顆粒層(inner nuclear layer)である。ただし、内顆粒層が完成するのは胎生の末期であり、それまでは外神経芽細胞層から内神経芽細胞層への幼若神経細胞の遊出が続く。 外神経芽細胞層は、このように多数の幼若神経細胞を送り出した後も、なお紡錘形の核が数列密に並んだ層として存続する。これらの細胞は最終的には光を感知する細胞、すなわち、錐体細胞(cone cells)および杆体細胞(rod cells)に分化するのであるが、これらの細胞の分化は網膜の構成要素のなかで最も遅い。 これらの細胞は、はじめは脳室上衣細胞と同じく、視室の中に繊毛を出している。胎生第6月になるとこの繊毛は消失し、これにかわって太い原形質性の突起が外境界膜を越えて視室の中に突出してくる。これが錐体(cone)および杆体(rod)である。錐体も杆体も、近位の内節と遠位の外節からできているが、光を感知して神経性興奮を起すのは外節である。外節は遠位に伸びて、色素上皮層の表面に達し、色素上皮細胞の微絨毛性の突起と嵌合して終わる。 眼杯の成立経過から明らかなように、眼杯の内腔、すなわち、視室に面している眼杯内板の表面と眼杯外板の表面は、共に脳室に接する脳の実質の内面に相当する。脳では内面を縁取る細胞は、表面から脳室に向って繊毛を送り出している。眼杯外板(網膜色素上皮層)では、この繊毛はメラニン顆粒を密に含む微絨毛様の突起となっている。眼杯内板ではこの微絨毛は消失し、これにかわって光を感知する錐体と杆体とが視室の中に突出してくる。そしてそれらの遠位端部が外節として伸びてきて、網膜色素上皮細胞の表面の微絨毛様の突起と嵌合するのである。したがって、網膜(眼杯内板)と色素上皮層(眼杯外板)の結合は緩やかであり、強固ではない。 錐体および杆体を出した残りの細胞質は、1本の細い繊維状の突起となって内方に伸び、その末端は小さい膨らみとなって、双極神経細胞の末梢性突起と接続する。双極神経細胞の中枢性突起は内方に進み、視神経細胞層の神経細胞の、主として樹状突起に接続する。 網膜におけるもう一つの構成要素である神経膠細胞(グリア細胞)も、胚芽層(外神経芽細胞層)から作り出される。すなわち、幼若神経細胞が盛んに作り出されている胎生第6週から第10週にかけて、グリア細胞および網膜に特有の支持細胞であるミュラー細胞(Mueller cells)も作り出される。ミュラー細胞は核を内神経芽細胞層(後の内顆粒層)に置く細長い柱状の細胞で、網膜の全層を貫き、網膜の内面と外面に、隣接する細胞の末端どうしが結合することによって、それぞれ内境界膜と外境界膜(inner and outer limiting membrane)を形成する。錐体および杆体は、この外境界膜を貫いて、視室の中に突出している。 このような網膜の組織発生は、すべて、まず眼杯の後極付近で始まり、ここから徐々に眼杯の前縁にむかって進んでいき、胎生第6月の終りごろになると、一応網膜視部の全領域で、全ての層が認められるようになる。しかし、網膜の最終的な完成はずっと遅く、部分的には出生後においてはじめて完成する。 |
| L. 視神経 |
| 視神経は、網膜の視神経細胞層の神経細胞から出た神経線維が眼杯茎の中に進入していくことによって成立する。「B. 眼裂およびその閉鎖」の項で述べたように、眼杯茎の遠位端部は、胎生裂の成立によって二重壁の管となっており、その内板は眼杯内板に直接続いている。視神経細胞層の神経細胞からでた軸索(視神経線維)は、網膜の最表層(縁帯)に入って内面に平行に走り、視神経繊維層を形成し、網膜の全領域から、眼杯茎に向って集まってきて、眼杯茎内板に進入し、眼杯茎内板およびその後方に続く眼杯茎を高度に肥厚させる。その結果、眼杯茎は閉鎖され、さらに眼裂も閉じるので、眼杯茎の遠位部は、中軸部に網膜中心動静脈を含む充実性の神経索となる。眼杯茎の近位部においても、視神経繊維の進入によって、内腔が消失して充実性となり、全体として視神経となる。 視神経は、脳の間葉性被膜である視神経鞘で包まれている。視神経鞘のうち、硬膜鞘は眼球の後極において強膜に直接移行する。 |
| M. 眼瞼および涙腺 |
| 眼瞼(eyelid)は胎生第2月において、眼杯の上方と下方に生じる皮膚の「ひだ」として発生する。この上下の眼瞼原基は、角膜原基の前方を被いながら速やかに発育して、胎生第4月になると、その自由縁が互いに接するようになり、ついで相癒着する。この癒着を眼瞼縫合という。これから約3ヶ月間胎児は眼を閉じた状態で成長する。この上下の眼瞼の癒着は眼瞼原基の上皮層に限られる。眼瞼の縁に沿って発生する睫毛(まつげ)および腺は、上下の眼瞼に共通の上皮層から分化し始める。瞼板腺(tarsal
gland)は眼瞼縁後部の上皮から発生する。瞼板(tarsal plate)はこの腺を取り巻く間葉組織の濃縮として形成される。胎生第6月中に眼瞼縫合は緩み始め、第7月において上下の眼瞼は再び分離する。 眼瞼の前面の上皮は、体の他の部分と同様に重層扁平上皮となるが、後面の上皮は重層円柱上皮となり、これを裏打ちする結合組織とともに結膜(conjunc-tiva)を形成する。眼瞼結膜は眼瞼の上縁と下縁で反転して、眼球結膜となって強膜の前面を被い、強膜溝のところで角膜上皮に移行する。眼瞼結膜が眼球結膜に反転する部分を結膜円蓋といい、両者に囲まれた腔を結膜嚢という。 涙腺(lacrimal gland)は、胎生第3月において上結膜円蓋の外側部から生じる7 ~10 本の上皮細胞索として発生する。涙を結膜嚢から鼻腔に導く鼻涙管などの涙道は、顔面が形成される時に、上顎突起と下顎突起の間に一過性に生じる溝(鼻涙溝)の底の上皮から形成される。 2011.11.08. 11.09. |
| 戻る |